


Информационная феноменология жизни. Часть I: Внутриклеточные информационные отношения
Азотистые основания:
Пурины: A – аденин; G – гуанин.
Пиримидины: С – цитозин; Т – тимин (для ДНК) или U – урацил (для РНК).
Комплиментарность пар пурин-пиримидиновых оснований ДНК: A(T; G(C.
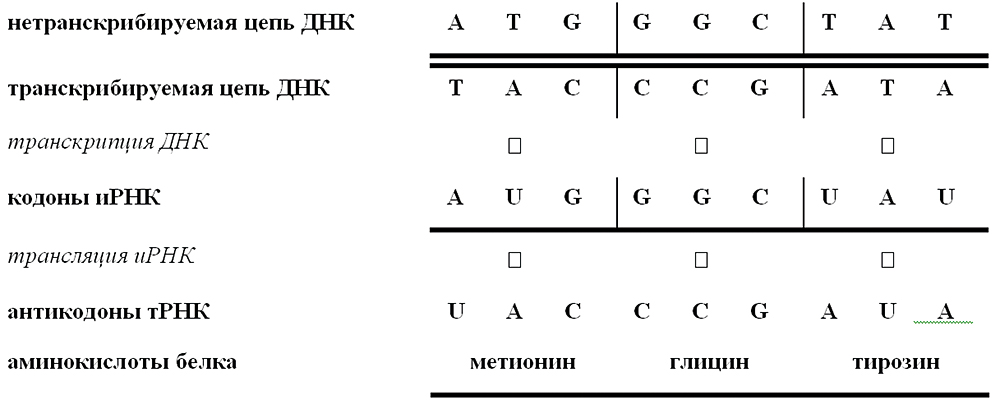
Подчеркнем, что на основе структуризации программной информации в реальных условиях исполнения программ обеспечивается возможность управления активизацией различных фрагментов (модулей) программ с учетом уже полученных результатов. Это в полной мере относится и к генетической информации. Активность генов, не только индуцибельных, но и конститутивных, и регуляторных, определяется объемом синтезированных генопродуктов – РНК и белков. Степень активности генов называется их экспрессией. Именно регуляцией активности генов объясняется, например, тот факт, что, несмотря на идентичность генотипов клеток многоклеточного организма, они значительно различаются по строению и функции. Переключение синтеза с одних белков на другие лежит в основе всякого развития, будь то репродукция вирусов, рост и спорообразование у бактерий, дифференцировка тканей и развитие эмбрионов у многоклеточных организмов. На каждом этапе таких процессов происходит активизация необходимых модулей генетической программы, обеспечивающих формирование фенотипической информации, которая, в свою очередь, обслуживает процессы метаболизма и развития клетки на каждом этапе ее жизни.
Прокариотная биосфера
Воплощение концепции модульной организации генетической программы прокариотных организмов, по-видимому, в значительной степени определило комбинаторный характер распределения признаков фенотипической системы бактерий. Одним из основных процессов поддержания такой направленности эволюции прокариот считается трансгенозис – перенос генов в форме фрагментов ДНК. Полагают, что в качестве исходного следует признать наличие в среде обитания живых существ большого числа всевозможных частиц, структур и обрывков ДНК и РНК. Кроме того, в среде обитания находится множество вирусов. Все эти содержащие генетическую информацию объединения представляют реальность в эволюции. Важным фактором при этом выступает отношение самих организмов к возможности интеграции чужеродных ДНК. Отмечается, что бактерии в этом плане обладают специальными приспособительными механизмами для выделения и для приобретения полноценного генетического материала извне. Здесь и трансдукция и трансформация. Одним из достаточно известных и изученных подобных процессов является обмен плазмидами.
Полагают, что такой генетический, информационный обмен играет особую роль у прокариот. Обмен генами между организмами, принадлежащими к далеким в систематическом отношении группам, открывает возможность приобретения сходных признаков не только на основе исторической общности происхождения или наследственности. Приспособление бактерий к использованию нового субстрата, определяющее принадлежность к соответствующей физиологической группе и соответственно к возможности занять определенную экологическую нишу, зависит от относительно небольшого числа ферментов. Объем генетической информации для их синтеза вполне совместим с механизмами трансгенозиса.
В прокариотных организмах сформировалась модульная структура программного обеспечения управления имманентными процессами развития клеток, обладающая вместе с тем определенными механизмами (интерфейсами) возможности восприятия внешних (трансцендентных) информационных воздействий, которые с учетом определенных критериев (барьеров различного уровня, включая кроме информационных всю совокупность имманентных биологических отношений) тем или иным способом включались в клеточный геном. Это обуславливало процессы коэволюции прокариотной биосферы в среде ее существования в различных ареалах верхних оболочек Земли.
Сформировавшиеся механизмы реализации внутриклеточных информационных отношений обеспечили возможности создания при их помощи широкого спектра соматических вариаций прокариотных организмов. Напомним еще раз: считается, что сложившиеся на этапе начала жизни механизмы и архитектура информационных отношений, обеспечивающих конвариантную редупликацию прокариотных организмов, не претерпели каких-либо значимых качественных изменений в ходе всей эволюции этого царства живой природы по настоящее время. Вместе с тем физиологическое разнообразие прокариот является фактом активных эволюционных процессов их физиолого-биохимической специализации. Не смотря на то, что эти процессы не касались непосредственно структуры сложившихся информационных отношений и не являются в связи с этим предметом по заявленной теме обсуждения, но их значимость в ресурсном обеспечении жизнедеятельности биологических организмов отнюдь не уступает значимости информационных отношений для развития биосферы в целом.
Если информационные отношения выступают имманентным модератором развития биологических организмов, то физиолого-биохимические аспекты обеспечивают их трансцендентные отношения с окружающей средой. Именно эта паритетность hard– и software и их адекватность составляют сущность биологических структур, реализуя процессы коэволюции в изменяющейся окружающей среде. Действительно, без необходимого ресурсного обеспечения бессмысленно говорить о возможностях реализации каких-либо информационных отношений. Это аксиоматическое свойство любых, в том числе и информационных, систем, к которым, как частный случай, относятся и биологические системы любого уровня. Заметим к тому же, что отсутствие информационного программного управления выводит любые физико-химические процессы из категории биологических. В связи с этим физиолого-биохимические факторы жизнедеятельности биологических организмов являются необходимым атрибутом возможности реализации основного свойства жизни – редупликации (тиражирования) за счет привлечения ресурсов внешней среды, которое обеспечивается действием имманентного информационно программного управления. Физиолого-биохимические аспекты биологических организмов определяют мощность их тиражирования в конкретных условиях и состояниях окружающей среды. Это, в свою очередь, создает условия формирования ареалов биосферы – среды, которая не только идентифицирует наличие жизни, но является необходимым условием, как ее существования в активном режиме, так и ее развития. Эти соображения являются поводом некоторого рассмотрения физиолого-биохимической эволюции прокариот и связанных с этим феноменологических аспектов развития жизни на нашей планете.
Бактерии на начальном этапе формирования биосферы были единственными живыми обитателями Земли, составляли и до сих пор составляют целостную и вполне самодостаточную систему в биосфере.
Вполне естественно предположение, которое высказал еще в 1909г. С.Орла-Йенсен (S.Orla-Jensen), что начальные формы прокариот были хемоавтотрофны. Определенным подтверждением этому являются реликтовые архебактерии. Анклавное распределение первичных прокариот создавало значительную дифференциацию условий, в которых реализовывался их жизненный цикл. Это привело к широкому спектру морфологического и физиологического разнообразия формирующихся типов бактерий. Вместе с тем, единообразие аппарата и механизмов действия информационных отношений, определяющих первичное (генетическое) программное управление онтогенезом прокариотных организмов, обеспечивало значительную совместимость филогенеза различных анклавов.
Анклавное распространение ареалов первичных прокариот было следствием широкого спектра различий окружающей среды, как в пространстве, так и во времени. То есть, находясь даже в конкретном анклаве, прокариотные организмы испытывали достаточно динамичные воздействия изменений окружающей среды в процессах филогенеза. Важным фактором при этом являлся углеродно-водородно-кислородный баланс в окружающей среде и доступность живым организмам веществ, необходимых для поддержания их циклов развития и размножения в сформировавшейся биосфере. Анклавный тип ареалов, с одной стороны, способствовал многообразию видов на основе разнообразия условий в различных анклавных ареалах, но, с другой стороны, ограничивал численность биоорганизмов, в связи с локальными ограничениями доступных ресурсов вещества в структуре каждого ареала. Кроме того, анклавный тип ареалов за счет специализации видов ограничивал возможности их распространения.
Естественно, что все эти ограничения достаточно жестко действовали в условиях существования только хемоавтотрофных форм прокариот. Ситуация значительным образом изменилась, когда в структуре цитоплазматической мембраны (ЦПМ) прокариот начали функционировать фоторецепторы. У первичных прокариот концентрация результатов синтеза белков – фенотипической информации, происходила главным образом в ЦПМ. Это обусловило активную роль ЦПМ не только в функции обеспечения границы клетки с окружающей средой, но и в функции своего рода клеточной органеллы, которая реализовывала воздействие фенотипической информации на жизнь клетки. Такая активность естественно сопровождалась достаточно высокой интенсивностью динамических преобразований ЦПМ, на которых «опробовались» новые функции, как трансцендентного аспекта (интерфейсы взаимодействия клетки с внешней средой) так и имманентного аспекта (участие в организации обратной связи влияния синтезированных в клетке продуктов на экспрессию первичной генетической информации). Поэтому не удивительно, что именно в структуре ЦПМ, например, сформировались, конструкции фотосинтетических пигментов.
Этот процесс, по-видимому, был обусловлен так же тем, что по своей структуре органические фотосинтетические пигменты сходны с липидами, которые являются основой организации мембран. Двухстадийность процессов фотосинтеза в его современной интерпретации, разделенного на световые и темновые реакции, подсказывает, что закрепление функций фотосинтеза происходило в ходе некоторых этапов эволюции. Возможно, это было связано с климатической динамикой в истории Земли. Например, около 2,5 млрд. лет назад значительно уменьшилась тектоническая активность, что уменьшило дегазацию недр. Ускорился и вывод углекислого газа из атмосферы. В результате всего за сотню-полторы миллионов лет основные запасы углекислого газа были выведены из атмосферы, наступило первое в истории земли мощное оледенение, известное как гуронское. Оно продолжалось более сотни миллионов лет, и средняя температура на поверхности Земли на уровне моря в это время составляла менее 10 °С.
В этих условиях энергетическое обеспечение реакций хемосинтеза значительно ограничилось по сравнению с предыдущим периодом, когда средняя температура на уровне моря составляла около 50–70 °С. Возможно, этим объясняется определенная бедность предшествовавших гуронскому оледенению первичных прокариот белковыми продуктами и высокая ферментативная активность их РНК компонент. Вовлечение световых реакций в биохимический цикл клетки, обеспечило таким клеткам определенные преимущества по сравнению с хемотрофными организмами.
Роль световых реакций заключается в синтезе за счет энергии света молекул АТФ и НАДФ Н2, которые в свою очередь обеспечивают энергией большинство супрамолекулярных процессов, протекающих в клетках. Воздействие кванта света обеспечивает ионизацию молекулы пигмента с выделением свободного электрона, который запускает ряд окислительно-восстановительных реакций. Непременным результатом этих реакций является потребление водорода для формирования НАДФ Н2. Источником водорода является либо вода – Н2О (например, для цианобактерий), либо сероводород – Н2S (например, для серобактерий) и т.п. Обычно оставшееся вещество источника водорода выделяется из организма. У цианобактерий выделяется кислород, а у серобактерий, соответственно вместо кислорода, выделяется сера. Такие побочные свойства клеточного метаболизма обуславливают накопление во внешней среде продуктов клеточного метаболизма. В частности с этим, обычно, связывается насыщение земной атмосферы кислородом.
Повышенная энергонасыщенность ЦПМ прокариотных клеток в результате световых реакций (энергетический метаболизм) создала возможности реализации на этой основе реакций конструктивного метаболизма. Учитывая, что это происходит в ЦПМ – пограничной клеточной оболочке непосредственно контактирующей с внешней средой, достаточно естественно, что в процессы изменения метаболизма на этом уровне были вовлечены как ассортимент питательных веществ из внешнего субстрата, так и интерфейсы трансцендентных отношений клетки с окружающей средой.
Появление для отдельных клеток нового источника энергии обеспечения жизненных процессов создало принципиально новую нишу жизненного пространства прокариотного мира. Создалась кажущиеся ситуация отсутствия конкуренции в ресурсопотреблении среды обитания. Но опыт показывает, что в анклавных условиях обычно наблюдаются четыре фазы развития популяции, включая: лиг-фазу – адаптацию бактерий в среде обитания, логарифмическую фазу – экспоненциального роста численности бактерий, стационарную фазу – динамического баланса гибели и образования новых клеток, фазу ускорения гибели – резкое сокращение размножения бактерий. Реализация таких сценариев активно ограничивает существование живых организмов не только внутри анклавов, но и по совокупности анклавов в целом. То есть должны существовать механизмы, регулирующие интенсивность роста численности живых организмов, обеспечивающие поддержание стационарной фазы, в сложившихся условиях экологической ниши для каждого вида живых организмов.
В условиях нашей биосферы такие механизмы в общем виде ее существования обеспечиваются разделением всех живых существ на экологические категории продуцентов, консументов и редуцентов (деструкторы). Динамическая обусловленность существования этих категорий живых организмов создает возможность сбалансированного круговорота веществ непосредственно в живых организмах и в среде их обитания. С феноменологической точки зрения можно отметить, что это отражает совокупное влияние имманентных и трансцендентных факторов на существование и развитие живых организмов, как самостоятельных развивающихся открытых систем.
Выше указывалось, что трансцендентное воздействие и интеграция в генетические файлы прокариот элементов ДНК и РНК из среды обитания в значительной степени определили комбинаторный характер распределения признаков для фенотипической системы бактерий. Этот факт феноменологически показывает взаимообусловленность эволюции прокариот как имманентными, так и трансцендентными факторами, подчеркивая значимость коэволюции в общем процессе формирования прокариотной биосферы. С таких же позиций коэволюции возможно представить процесс расслоения прокариотных организмов на автотрофов и гетеротрофов. Отметим, что это не «отбор» (естественный, либо иной), а именно процесс коэволюции, развивающийся на основе синергетических отношений имманентной изменчивости организмов и, на основе этого, их трансцендентного воздействия, в свою очередь, на изменчивость среды обитания и восприятия этой изменчивости.
В рамках искусственных информационных систем перманентная модернизация аппаратных средств и программного обеспечения в соответствии со складывающимися условиями их применения и возможностями эксплуатации получила название стратегии Up-grade. Можно заметить, что с позиций феноменологии такой процесс в значительной степени коррелируется по существу совершенствования информационных систем с процессами коэволюции развития прокариотной биосферы.
В процессе формирования прокариотной биосферы на основе многообразия групп бактерий и их типов внутри групп фактически были заложены указанные три экологические категории живых организмов: продуценты, консументы и редуценты (деструкторы). Наличие этих категорий создает возможности динамического круговорота «живой материи» в природе. Это особенно существенно при возникновении ограничений либо трансформаций каких-либо ресурсов внешней среды. Такие ситуации могут возникать не только при непосредственном обеднении среды по каким-либо ресурсным показателям, но и при изменении их соотношений. В какой-то степени, ярким примером таких изменений является формирование современной атмосферы как результата жизнедеятельности прокариотной биосферы.
Категория продуцентов, объединяющая автотрофные организмы, обеспечивает биосферу «живой материей» как для собственного самовоспроизводства, так и для функционирования гетеротрофных категорий живых организмов. Использование термина «живая материя» имеет серьезное обоснование. Суть его в том, что базовые супрамолекулярные структуры живых клеток образованы на определенной упорядоченности энантиомеров химических соединений. Это создает хиральность физико-химической среды транспортных уровней реализации информационных отношений, принципиально связанных с необходимостью жесткого соответствия логической структуре программного обеспечения развития живых клеток на основе как генетической, так и фенотипической информации. Таким образом, содержательной стороной круговорота «живой материи» является поддержание определенной концентрации в биосфере хиральных продуктов, на основе которых возможно создание средств для реализации живыми организмами необходимых имманентных программ их развития и существования. Это достигается за счет эксклюзивного индивидуального синтеза каждой молекулы энантиомера в клетках живых организмов на основе программного управления, реализуемого с помощью генетической и фенотипической информации, хранимой и создаваемой в клетках. Продуценты в этой цепи являются звеном первичного преобразования костного вещества внешней среды в элементы «живой материи».
Исходным генератором (основой) этих преобразований является жизнедеятельность цианобактерий (сине-зеленые водоросли). Необходимо отметить, что и в настоящее время они играют доминирующую роль в обеспечении углеродно-кислодродного баланса на поверхности земного шара и в его атмосфере.
Важным условием, обеспечивающим поддержание стационарной фазы динамического баланса гибели и образования новых клеток, является регулирование интенсивности роста численности живых организмов в сложившихся условиях экологических ниш. Процесс расслоения прокариотных организмов на автотрофов и гетеротрофов породил формирование механизма этого регулирования с участием консументов и редуцентов (деструкторов). Консументы, в общем случае, являются регуляторами численности продуцентов, обеспечивая согласованность интенсивности инициализации «живой материи» объективному ресурсному обеспечению в окружающей среде. Однако такое простое регулирование в чистом виде могло привести к обеднению среды обитания костным веществом для функционирования продуцентов. Утилизация гибнущих организмов и продуктов их жизнедеятельности обеспечивается редуцентами (деструкторами). За счет этой утилизации омертвевшая часть живой материи после разложения ее до уровня неорганических соединений вновь возвращается как сырье для нового цикла продолжения Жизни.
С учетом представлений о развитии биосферы на Земле, созданные в ходе эволюции прокариот механизмы их адаптации (естественная реализация стратегии Up-grade) в условиях изменчивости среды обитания и регулирования численности в соответствии с реалиями ресурсного обеспечения достаточно убедительно показывают высокий уровень самодостаточности царства прокариотных организмов.
Эукариотные клетки (Eucaryota)
На общедоступном языке мы можем назвать ядро АДМИНИСТРАТОРОМ КЛЕТКИ. Две главные черты роднят его с наиболее известными администраторами: оно стремиться плодить себе подобных и успешно отражает все наши попытки узнать, чем же именно оно занимается. Только попытавшись обойтись без него, мы узнаём, наконец, что оно действительно работает.
Д. МэйзиЖивая природа на Земле, существующая в настоящее время, представлена двумя классами внутриклеточной организации живых организмов: прокариоты (Procaryota) и эукариоты (Eucaryota).
В научном представлении общепризнанно, что прокариоты считаются родоначальниками нашей жизни на планете. Сформировав почти 4млд лет назад первичную биосферу, прокариоты и в настоящее время в значительной степени определяют динамику круговорота живой материи на Земле. В этом процессе только прокариоты, например, обеспечивают биосферу поступлением ряда веществ из «костной» (абиогенной) материи (азот, фосфор и др.), которые затем активно включаются в формирование органических соединений.
Феноменологическая значимость прокариот в жизни современной биосферы и планеты в целом отмечена таким глобальным явлением, например, как образование кислородной атмосферы Земли. В рамках современных представлений, основную роль в этом сыграли цианобактерии (сине-зеленые водоросли в иной терминологии). И в настоящее время, не смотря на огромную биомассу зеленых растений, влияние цианобактерий в динамике углеродно-кислородного круговорота на планете остается весьма значимым, а возможно и доминирующим. Биомасса микроорганизмов океана составляет около трети всей биомассы биоты планеты, биомасса бактерий суши сравнима с биомассой растений. Таким образом, считается, что биомасса прокариот – от половины до 90 % всего живого вещества биосферы. Не удивительно поэтому, почему так высока степень влияния прокариот практически по всем элементам биохимических циклов живой природы.
Достаточно интересной, кроме того, является гипотеза биогенного формирования не только атмосферы планеты, но и океанов на поверхности Земли. Гипотезы о появлении гидросферы на нашей планете, представляющие генерацию воды только как неорганический процесс, имеют пока что много необъяснимых проблем. В большинстве своем они связаны с временными последовательностями заполнения Мирового океана. Исследования по распространению и жизненным циклам водородных бактерий в природе, проведенные М. И.Беляевой еще в 1950-ых годах в Казанском университете, вполне убедительно показывают возможность существенного влияния живой природы на гидросферу планеты не только в плане регулирования водостоков, но и в интенсивности накопления водных ресурсов на основе биологического синтеза воды из абиогенной материи планеты.
Более того, по истории осадочных оболочек Земли накоплен достаточный материал, на основе анализа которого уверенно утверждается, что органического углерода всегда было много и некоторые древние породы сильно графитизированы. В этом случае вполне сильна и аргументирована гипотеза, что органический углерод за 3,5 млрд. лет существования осадочных пород на Земле образовался путем автотрофной ассимиляции углекислоты. Последний вывод основан на том, что включение СО2 в обмен под действием рибулозобисфосфаткарбоксилазы приводит к облегчению изотопного состава органического углерода13С/12С = – 25‰. Именно такое облегчение наблюдается на всем протяжении осадочной летописи. Поскольку иные способы ассимиляции СО2, например анаэробами, дают иной изотопный состав – то утверждается, что органический углерод во все это время синтезировался таким же путем, как сейчас.
Академик Г. А.Заварзин так характеризует процессы коэволюции прокариотных организмов. «С первого момента их регистрации на Земле 3,5 млрд. лет назад до настоящего времени бактерии действуют как полноценная трофическая система, составленная разнородными компонентами… Эволюция их сообществ более всего напоминает сукцессию [38], когда массовое развитие, вне зависимости от времени появления одиночных представителей определяется изменением условий существования, в том числе происходящих и под воздействием самих организмов. Во время деятельности прокариотной системы на Земле происходит однонаправленное изменение физико-химических условий, лучше всего иллюстрируемое эпохой железорудных формаций. Так, например, судя по изотопии серы, полный серный цикл с восстановлением сульфатов начал работать около 2 млрд. лет назад, что связано с распространением определенных групп бактерий, но в меньшем масштабе такой процесс мог идти и ранее, о чем свидетельствуют сульфаты древних эвапоритов и строматолитов. Разумеется, такая постановка вопроса о доминировании сукцессии сообществ над появлением новых видов не снимает возможности микроэволюции бактерий. Она отчетливо наблюдается, например, под воздействием антропогенных факторов при распространении резистентных форм, причем здесь участвуют специфические для бактерий механизмы. Представление о сукцессии бактериальных сообществ ставит вопрос о персистентности [39] видов бактерий. На этот вопрос трудно ответить в общем виде».
В этой связи достаточно интересны результаты работ датского ученого Тома Фенчеля о распространенности и значимости тиобиоса[40] в настоящее время. Буквально под ногами, в прибрежных илистых осадках на глубине чуть более пяти сантиметров, им обнаружено огромное разнообразие многоклеточных животных, живущих… в сероводородной среде. Стало очевидно, что и в настоящее время животный мир на Земле представляется, по крайней мере, в виде двух частей: одна дышит кислородом (оксибиос), другая – сероводородом (тиобиос). Не исключено, что именно сероводородные бактерии когда-то способствовали началу эволюции многоклеточных животных. Сероводородные бактерии – древнейшая форма жизни на планете, развивавшаяся параллельно и независимо от кислородной жизни. Это новая гипотеза о соотношении развития различных форм биологической жизни на нашей планете, которая, по мнению биологов, нуждается в серьезном научном исследовании.